La « naturalité » d'un espace est une notion intégratrice qui « synthétise l'expression en un lieu des propriétés écologiques intrinsèques de la Nature » (VALLAURI et al. 2017). Cette notion fait fortement écho avec les forêts qui constituent la végétation spontanée et ultime sur la quasi-totalité du territoire du Massif central (à l'exception des stations les plus sèches ou humides, ainsi que des crêtes ventées des plus hauts sommets). Elle regroupe les caractéristiques propres aux forêts naturelles : diversité des espèces, indigénat de celles-ci, complexité structurale et maturité des peuplements, microhabitats, nécromasse, intégrité des processus dynamiques et fonctionnels assurant notamment le renouvellement et la résilience des peuplements face aux perturbations, continuité spatiale et temporelle (ancienneté)…


Ces éléments se retrouvent déjà dans les concepts « d'état de conservation » ou dans « l'indice de biodiversité potentielle » mais la « naturalité » intègre également des valeurs plus philosophiques et éthiques comme le « sentiment de nature » tout en tenant compte des traces d'activités anthropiques, mais aussi de la « spontanéité de la dynamique observée ».
Pour autant, si la biodiversité est très élevée dans les écosystèmes forestiers à forte naturalité, dont l'état de conservation est également très bon, les forêts marquées par une anthropisation actuelle ou passée ne sont pas toujours synonymes de faible biodiversité pour autant. Certains microhabitats actuellement très favorables à la biodiversité sont finalement issus de pratiques anciennes abandonnées ! On pensera, par exemple, aux blessures et nécroses causées par la coupe des cépées et arbres têtards (anciens arbres ruraux ou arbres forestiers dont les rameaux feuillés étaient récoltés pour nourrir le bétail) permettant plus tard l'accueil d'une faune cavernicole exceptionnelle…
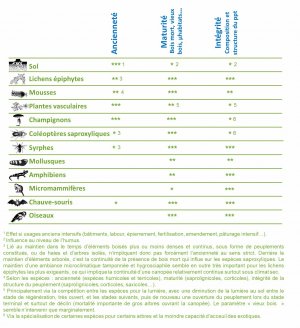
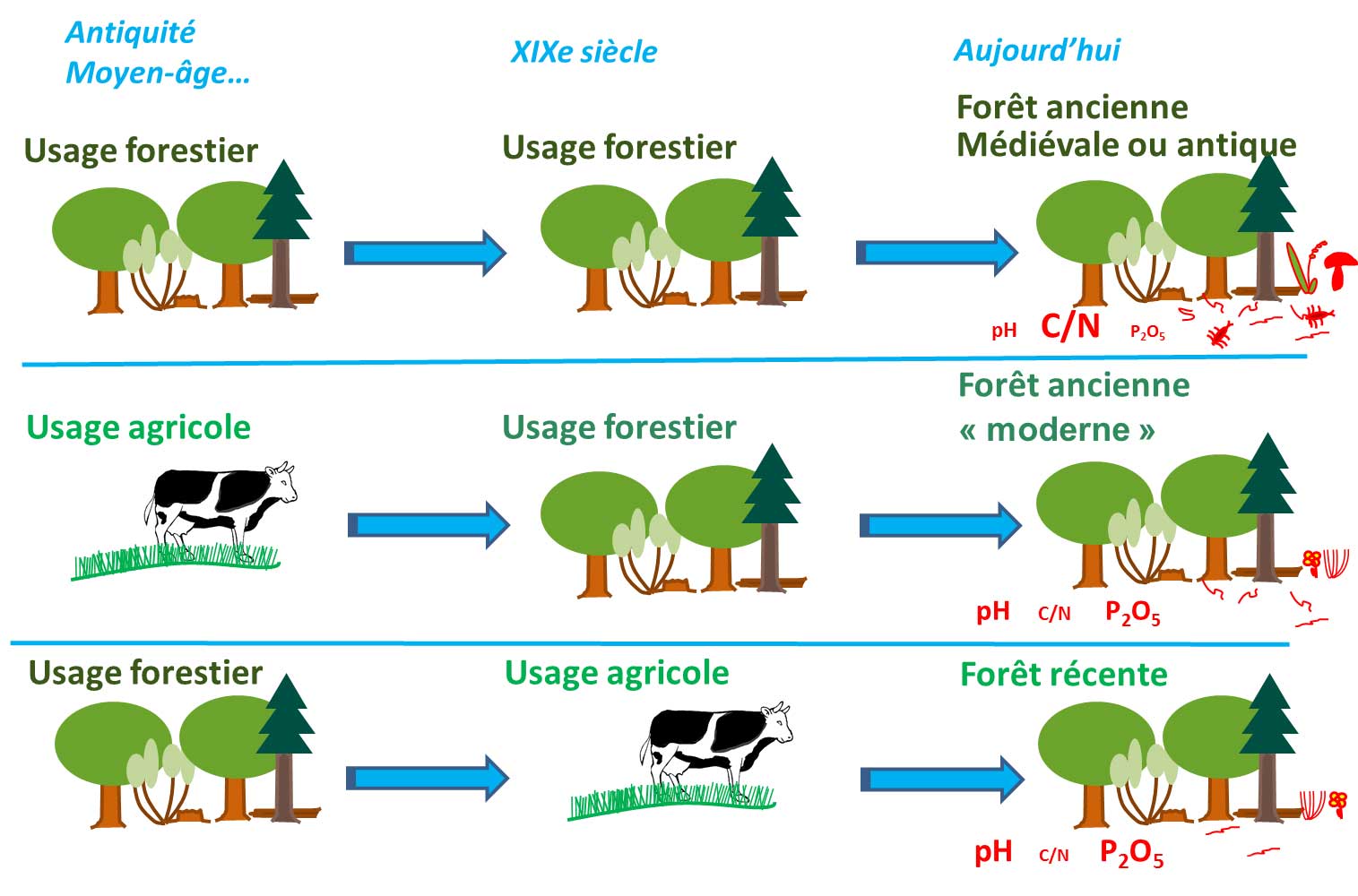
Les forêts anciennes se distinguent par l'absence de défrichement depuis au moins la première moitié du XIXème siècle, quelle que soit la gestion forestière pratiquée. L'ancienneté est un gradient, et cette continuité forestière peut remonter à des temps beaucoup plus anciens (forêt médiévale, forêt antique…). Ainsi 20 à 40 % des forêts actuelles seraient anciennes, avec des valeurs plus extrêmes (entre à peine 10% et plus de 80 %) et de fortes disparités en fonction des territoires.
En raison de leur histoire, ces forêts peuvent constituer des hauts-lieux de la biodiversité en hébergeant des espèces qui ne se rencontrent que rarement ailleurs. Lorsqu'une forêt est défrichée (agriculture, urbanisation, routes, etc.), on peut observer la disparition d'un grand nombre d'espèces forestières, mais aussi des changements profonds et durables dans le sol.
Même après reconstitution du couvert arboré, les bouleversements dus au pâturage, à la fertilisation, au chaulage, à l'épierrement ou au labour influent sur la flore, la faune et les champignons du sol et du sous-bois durant des siècles. Dès lors, après une période d'usage non forestier, le retour des espèces typiquement forestières peut s'avérer particulièrement long. C'est notamment le cas des plantes à faible capacité de dispersion qui, contrairement à certaines plantes aux graines voyageuses ou à des espèces animales très mobiles, mettront des siècles à conquérir de nouveaux terrains. Certaines plantes à bulbe ou à rhizome (Muguet, Maïanthème à deux feuilles, Sceau de Salomon multiflore…), ou des plantes dont les graines sont disséminées par les fourmis (Anémone sylvie, Euphorbes, Laîche des bois, Luzule des bois, Mélique uniflore…) se déplacent de moins de 30 mètres par siècle !



Dans le massif forestier de Tronçais (Allier), 1500 ans après le retour de la forêt, à l'emplacement d'anciennes fermes gallo-romaines, des différences s'observent entre les secteurs demeurés en forêts ou voire en landes et ceux qui ont été cultivés !
L'ancienneté joue donc un rôle prépondérant sur la biodiversité présente et on comprend rapidement l'intérêt que portent les écologues à l'identification des forêts les plus anciennes. Si les études diachroniques actuelles permettent de qualifier les forêts qui l'étaient déjà il y a 2 siècles, les études montrent qu'il serait encore plus judicieux de caractériser celles qui le sont depuis des temps beaucoup plus anciens, peut-être même depuis la reconquête de la forêt après la dernière glaciation !
Toutes les espèces forestières ne sont pas nécessairement liées à l'ancienneté. Davantage semblent dépendre d'éléments liés à la maturité, comme la présence de vieux arbres, souvent porteurs de microhabitats, de divers types de bois mort, debout ou au sol, ou encore d'arbres de fort diamètre. Un tiers à un quart des espèces forestières dépendent ainsi du bois mort, et de nombreuses espèces vivent sur les troncs et dans les cavités. Certaines espèces cavicoles et saproxyliques nécessitent la présence de 20 à 50 m3/ha de bois mort pour se maintenir, notamment sous la forme de gros arbres (chablis ou chandelles). Les plus sensibles réclament plus de 100 m3/ha, volumes couramment observés dans les forêts naturelles.
Si l'on pense naturellement aux oiseaux (chouettes, pics, mésanges), au mammifères (chauves-souris, écureuils, martre…) ou encore aux insectes (coléoptères, fourmis…), le bois mort ou pourrissant est aussi un support indispensable à de nombreuses espèces de plantes dont certaines bryophytes rares et remarquable comme la Buxbaumie verte et bien évidemment, bon nombre de lichens et champignons.
Ainsi dans une forêt ancienne, la biodiversité est d'autant plus remarquable que la gestion actuelle et passée a ménagé la présence d'arbres sénescents, porteurs de microhabitats, ainsi que de bois mort, éléments indispensables à de nombreuses espèces forestières. La pratique puis l'abandon de certains modes de gestion, comme le taillis ou des tailles de type « arbre têtard », ont pu favoriser certains microhabitats et une biodiversité remarquable, y compris dans les peuplements les plus gérés.





Mais c'est dans les peuplements dans lesquels aucune coupe n'a été réalisée depuis parfois plus d'un demi-siècle que le potentiel biologique est le plus élevé, surtout pour les groupes taxonomiques les plus sensibles au dérangement ou à la mise en lumière. Ainsi, que la forêt continue d'être exploitée ou non, la biodiversité est d'autant plus remarquable que la forêt est ancienne, qu'elle ait conservé dans le temps des attributs de forêt naturelles (essences autochtones notamment celles du « climax », structure étagée et hétérogène…), et que le compartiment « vieux bois » n'ait jamais disparu.
Les peuplements ayant dépassé l'âge d'exploitabilité économique, âgés ou en cours de maturation biologique, ne représenteraient en France que 2% des forêts de chênes et 5 % des forêts de hêtres d'après les données de l'Inventaire forestier national. En l'absence d'intervention depuis au moins 50 ans, on peut parfois parler de « forêt à caractère naturel », ou encore de « forêt subnaturelle » ou « vieille forêt ». Ce sont les plus rares et les plus précieuses pour la biodiversité mais elles ne représenteraient que 0,2 % des forêts de France métropolitaine (MAAPRAT-IFN 2011).
Dans un contexte d'intensification de l'exploitation forestière, il paraît dès lors crucial d'identifier les peuplements les plus remarquables et âgés, ce que permet la méthode développée par le CBN Massif central et ses partenaires.
À l'échelle du peuplement, la maturité est une notion qui dépasse le compartiment « vieux-bois » (arbres morts et âgés, dendromicrohabitats, etc.). Elle se traduit au niveau du sol, notamment de la litière, par l'accumulation progressive de matière organique (feuilles, bois mort…) au fil des années avant d'être totalement ou partiellement minéralisée en cas de coupe ou de trouée de taille importante, consécutive à une tempête par exemple.


La structuration verticale en plusieurs strates de végétation et son hétérogénéité spatiale (mosaïque de peuplements ou bouquets d'âge différents, présence de petites trouées, alternance spatiale et temporelle des essences principales…) sont globalement favorables à la biodiversité typiquement forestière, et certains groupes y sont très sensibles (vertébrés notamment). Ces caractéristiques se retrouvent dans les forêts à caractère naturel, mais aussi dans les peuplements irréguliers, dans lesquels le forestier aura privilégié la coupe d'une partie des arbres à leur récolte complète (coupe dite « à blanc »).
Dans le Massif central, à l'exclusion du Châtaignier considéré comme naturalisé (au moins depuis le moyen-âge dans plusieurs secteurs) et du Pin à crochet introduit dans de nombreuses tourbières (mais probablement autochtone dans plusieurs tourbières du Haut-Forez et des Monts Dore), les botanistes comptent 37 espèces d'arbres originaires du Massif central et de nombreux hydrides.
La diversité en essences autochtones est globalement favorable à la biodiversité mais elle ne s'exprime pas de la même manière selon que la forêt soit modifiée par l'homme, selon le stade d'évolution considéré (clairière > recolonisation > peuplement mâture…) ou encore les conditions écologiques en place.
En montagne et en conditions naturelles, par exemple, hormis les stations à forte contraintes (humides, très sèches, sur éboulis ou blocs, sur sol très acide…), un mélange de Hêtre et de Sapin blanc, en proportion variable, domine la plupart du temps. En plaine, les hêtres sont naturellement accompagnés par les chênes tandis que ces derniers les remplacent totalement en zone méditerranéenne.
Parmi la fonge, la faune et les mousses, certaines espèces très spécialisées sont étroitement associées aux essences forestières pionnières (notamment les bouleaux et saules), rencontrées dans les coupes, accrus et sur les stations à forte contrainte. Une bonne partie de la biodiversité s'est, quant à elle, adaptée aux essences dominantes caractérisant le « climax », autrement dit le stade ultime d'évolution de dynamique forestière en l'absence de perturbations humaines.


Dans le Massif central tout comme la majorité des régions françaises, la sylviculture a néanmoins favorisé l'une des espèces caractéristiques des stades climaciques au détriment des autres espèces naturellement dominantes. On parle alors de « sylvofaciès ». C'est le cas du Hêtre lorsque le taillis a été pratiqué en montagne ou, au contraire, du Sapin sélectionné alors pour son bois d'œuvre. En plaine, ce sont davantage les chênes qui ont été sélectionnés pour leur bois d'œuvre, notamment pour réaliser les tonneaux et anciennement des navires.
Les forêts du Massif central comptent aujourd'hui une quarantaine d'espèces exotiques dont principalement l'Épicéa commun (originaire des Alpes, Carpates, Europe de l'Est et du Nord), le Douglas (côte ouest d'Amérique du Nord), le Robinier faux-acacia, divers peupliers américains ou hybrides. De nombreuses autres espèces peuvent se rencontrer, à la faveur de plantations (Mélèze d'Europe, Chêne rouge d'Amérique, plusieurs espèces de pins, épicéas ou sapins nord-américains, Pin noir, etc.), ou échappés de parcs et de jardins. Ces espèces peuvent être moins adaptées au territoire, surtout si elles viennent d'un autre continent, et se montrent surtout moins favorables à la biodiversité d'autant plus lorsqu'elles s'avèrent envahissantes (cas du Robinier, de l'Érable negundo ou de l'Ailantes en forêts alluviales), ou lorsqu'elles sont plantées en peuplements équiens mono spécifiques, a plus forte raison s'ils n'ont pas été suffisamment d'éclaircis.

En France, au fil des défrichements successifs, les forêts ont progressivement été fragmentées en plusieurs petits espaces boisés séparés les uns des autres par des territoires « ouverts », parfois sur de grande distance. Dans ce contexte d'isolation progressive de leurs populations, la survie des espèces qu'elles soient animales ou végétales, dépend étroitement de leur capacité à assurer un brassage génétique suffisant. Les domaines vitaux de nombreuses espèces forestières étant souvent vastes (notamment parmi les vertébrés supérieurs), elles ne sauraient subsister qu'au sein de quelques petits ilots anciens.
Si les forêts anciennes et les peuplements matures constituent des réservoirs de biodiversité, les forêts récentes ainsi que le maillage bocager contribuent à la (re-)connexion boisée entre ces massifs anciens. Ils permettent ainsi la migration d'espèces dans un contexte de reconquête des massifs récents à partir des forêts anciennes (notamment depuis la fin du XIXème siècle), et plus récemment dans un contexte d'adaptation de la biodiversité aux changements climatiques.
On comprend alors qu'au-delà de la prise en compte de la fonctionnalité des écosystèmes forestiers et de la continuité temporelle évoquée précédemment (ancienneté, maturité, structure ou composition des peuplements), la sauvegarde des réservoirs de biodiversité que constituent les forêts anciennes et matures nécessite une certaine continuité spatiale. Une réflexion sur la connectivité et la perméabilité de la trame forestière dans son ensemble est donc nécessaire.
Au sud du territoire, la trame écologique du Massif central identifie notamment cet enjeu de continuité forestière à travers le maintien d'un axe forestier depuis l'Ardèche jusqu'à l'Aude assurant la connexion entre les Pyrénées et les Alpes, mais cet enjeu se pose en d'autres endroits et à d'autres échelles.
Outre sa répartition spatiale, la fonctionnalité de cette trame forestière dépend également de sa qualité et de son maintien dynamique au cours du temps. En son sein, la conservation d'une « trame de vieux bois », indispensable à la survie d'une grand nombre d'espèces, comprenant un maillage d'arbres morts, sénescents ou porteurs de microhabitats, d'ilots de sénescences et de zones plus vastes en libre évolution, constitue un enjeu fort.